Index
-
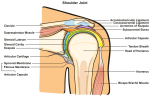
Motor Control considerations in the cervical spine
-
Muscle layers
-
Functional Synergies
-
Cervical Spine reflexes
-
Motor control of the trunk
-
Principles of Motor Control
-
Intrinsic Lumbo-pelvic muscles
-
Sensors
-
Controller
-
Lumbo-pelvic stability
-
Closed Loop control of the trunk
-
Motor Control - Long loop reflexes
-
Control of muscle stiffness
-
Effect of Pain and Injury on Motor Control
-
Pain Adaptation Model
-
Motor control – task conflict
-
Motor Control and Pain
-
Pain
-
Pain & the Brain
-
Reflexes
-
Muscle hyper/hypo-activity and chronic pain
-
Models of Motor Control
-
Nervous System Plasticity
-
Chronic Pain and Motor Control
-
Central Mechanisms of Chronic Pain
-
Anterior Cingulate Cortex (ACC)
-
Implication of neural changes
Motor Control considerations in the cervical spine
• Action cannot be considered as the sum of isolated movements
• Control operations are very much dependent upon the goal of the movement
• Cervical spine is not analogous to the rest of the spinal column due to it’s large degrees of freedom and specific inputs from intero- and extero-ceptors
• Issues of control must also consider the redundancies (spare capacity) within the system
• 20 pairs of muscles many of which can perform similar actions (Peterson et al 1989)
• Ultimate degrees of freedom problem is how to reduce/simplify the movement to be as efficient as possible (Bernstein 1967)
• Overall the number of independently controlled muscle elements (including compartmentalisation and subdivisions) exceeds the degree of freedom
• Many neck muscles have multiple insertions and multiple functions whose variability is task dependent (Richmond et al 1991, 1992)
• 8 joints with 6 degrees of freedom each – 3 rotational and 3 translational
• Simplified the greatest degrees of freedom occur for rotation at C1/2, flexion C1/2, and extension 0/C1, lateral bending across all vertebrae
• Head represents only 7% body weight but has 20 muscles linking the skull to the spine (Sherk & Parke 1983). The CNS programmes neck muscles to respond to specific directions of movement rather than generating an infinite variety of motor patterns (Keshner et al 1992)
• Neck muscle morphometry reveals heterogeneity w.r.t. musculotendon, sarcomere, fascicle length, angles of pennation, muscle fibre type (density of I vs II), and instead of distinct tendons the muscles attached to bone through an architecturally complex internal tendon & aponeurosis (Kamibayashi & Richmond 1998)
Muscle layers
• Muscles arranged in layers with the outer layer connecting the skull and shoulder girdle, with the deeper layers linking cervical and thoracic vertebrae.
• Superficial layer includes trapezius (whose fibres are sparse above C4) and sternocleidomastoid, the next layer include long dorsal (splenius capitis, semispinalis capitis and longissimus capitis) and long ventral (longus capitis), slightly deeper to this lie the splenius cervicis, semispinalis cervicis, longissimus cervicis and longus colli which link the vertebrae,
• whilst the deepest layers are the suboccipital muscles which include the rectus capitis major/minor, obliquus
capitus superior/inferior
Functional Synergies
• The problem of the controller is the optimisation between mobility and stability
• Muscle synergy has been described as “a consistent grouping of muscles to accomplish a specific task” (Turvey et al 1982) i.e muscles spanning several joints yet acting as a single unit (Buchanan et al 1989)
• Separate actions of 2 groups of muscle would assist the head-neck controller to meet the task of stability and movement (Thomsen et al 1994)
• Several muscles can achieve the same direction of movement. The controlled parameter appears to be the required force vector rather than the specific force lever arm of any one muscle (Macpherson 1988, 1991)
• In Rhesus monkeys during the upright position, muscle activation patterns were in functional groupings determined by their anatomical attachments, whereas in the quadrupedal position, gravity acting horizontally on the head produced greater activation and a collective response of the muscles suggesting a connection between central recruitment and the mechanical requirements of the task (Mayoux-Benhamou et al 1997)
• A mathematical model of head-neck control (Peng et al 1996) has identified stiffness and the vestibulocollic reflex as the primary contributors for the stabilisation of the head in space
• Vestibulocollic and cervicocollic reflexes respond reactively to acceleration and proprioceptive stimuli to orientate the head in space (VCR) maintain the head on trunk (CCR)
• Voluntary responses are those required for tracking and are based on exteroceptive information (visual, auditory, olfactory)
• Vestibulospinal pathways have monosynaptic and disynaptic descending inputs onto cervical motoneurones which have a distinct orienting function (Karlberg et al 1995).
• Other descending pathways, particular the reticulospinal pathways have been shown to be equally important in eliciting the VCR response (Peterson1984)
Cervical Spine reflexes
• The VCR produces counter-rotation of the head on trunk during postural disturbances
• The CCR stabilizes the head on the body and can provide information about the rotation of the head on the trunk
• Recent findings (Gdowski & McCrea 2000) suggest that correct alignment of the head with the trunk and with the gravitoinertial vertical (Imai et al 2001) requires that the vestibular system receive ascending somatosensory inputs (Keshner 2005)
• Semicircular canals are stimulated by angular acceleration in the horizontal plane
• The evidence suggests the presence of more signal processing than expected from a monosynaptic pathway, with evidence of a pre-synaptic inhibition in the CCR response (Banovetz et al 1995)
• The otoliths are stimulated by linear accelerations of the head and their inputs have been found to modify both eye and head stabilization (Schor et al 1985), however inputs from the otoliths cannot compensate perfectly to eye and neck responses (Borel & Lacour 1982)
• It was proposed that compensatory head pitch movements were produced predominantly by the angular vestibocollic reflex at slow walking speeds and by linear vestibulocollic reflex at high speeds (Hirasaki et al 1999)
• Sinusoidal stimulation in the horizontal and vertical planes (with the trunk fixed) indicate that at below 1Hz, the head is primarily stabilized by longer latency voluntary pathways observed as anticipatory torques in the neck muscles or responses controlled by visual signals, whereas at higher at frequencies, mechanical factors (e.g. inertia, stiffness and viscoelasticity) become important and act with little delay (Keshner 2005)
• Linear translations with the trunk free to move, revealed a significant somatosensory component to head stabilization (Vibert et al 2001)
Motor control of the trunk
• ‘internal model of body dynamics’ – comparative centre of input to expectations evaluating the response
• “Optimal trunk function is a complex interplay between movement and control of the integrity of the spine and pelvis at the intersegmental level, at a global level involving the control of orientation, and the contribution of the trunk to maintain equilibrium of the body w.r.t gravity and external forces” (Paul Hodges 2005)
• Even static postures involve movement with small cyclical movements occurring to counter respiration (Hodges et al 2002)
• Movement is used by the CNS to maintain stability and minimize energy expenditure e.g. small movements of the trunk prior to limb movements (Hodges et al 1999, 2000) and rotation of the pelvis around each orthogonal axis during gait (Perry 1992)
• Models of stability must incorporate 3 levels of control
– Intersegmental control - orientation control - control of equilibrium
• Euler model of stability using buckling failure (90N)
– Argues that the activity and stiffness of antagonistic muscles is required for lumbar spine equilibrium
– Considers muscles as ‘guy wires’ to stiffen the intervertebral joints that they span (Criso & Panjabi 1991)
– Static consideration whereby few investigations have analysed this in dynamic terms (Cholewicki et al 1997)
• Beyond ‘buckling’, spinal health must consider control of global and intersegmental motion and the contribution of the trunk to postural equilibrium
• Control of translation and rotation during an arc of movement (Bogduk et al 1995) must also be incorporated into a model of stability, where segmental control is an essential element (Crisco & Panjabi 1991)
• Control of body equilibrium during changes in the centre of mass particularly in situations where there is a conflict between spinal orientation and balance through movement of the trunk
Principles of Motor Control
• ‘neutral zone’ is the area around the neutral position where minimal stiffness occurs (Panjabi 1992)
• It has been argued that the ‘neutral zone’ increases in cases of chronic instability (Panjabi 1992)
• Control elements
– Sensors
– Control/planning system to analyse and prepare for execution
– Muscles to execute the response
• Muscle elements to consider include
– redundancy
– muscle architecture whereby muscles are biomechanically more suited to either motion or stability (Bergmark 1989; Richardson et al 1999, Sahrman 2002)
– muscle function can be divided into extremes in the spectrum as well as medium spectrum function for
• control of intervertebral motion,
• torque production
• spinal orientation
– Bergmark (1980) described muscles as either ‘global’ or ‘local’ based on anatomical characteristics
• Local muscles have limited ‘moment arm’ and cross one/few segments with an ideal anatomy to control
intervertebral movement e.g. multifidus, intertransversarii and interspinalis, transverse abdominis
• The global muscles have attachments to the pelvis and thorax and thus transcends multiple segments
which have larger moment arms and therefore larger torques and hence function to orient the trunk and
correct equilibrium e.g. erector spinae, lateral fibres of iliopsoas and quadratus lumborum, external and
internal obliques, rectus abdominis
• Synergy between local and global muscles is necessary for efficient movement. Excessive co-activation of
global muscles can control shear and rotation, it can also lead increased compressive load on the
intersegmental segments such as discs and joints
Intrinsic Lumbo-pelvic muscles
• Thoracolumbar fascia consists of 3 layers
– Posterior attaching to the spinous processes
– Middle layer attaching to the transverse processes
– Anterior layer crosses the quadratus lumborum (Williams et al 1989)
• Transverse Abdominis
– Role in the modulation of the intra-abdominal pressure (IAP) (Cresswell et al 1992, 1994) as well as
contribute to spinal stiffness (Hodges et al 2001)
– Fascial tension may directly restrict intervertebral motion or may provide gentle segmental compression through the posterior fibres of the thoracolumbar fascia (Gracovetsky et al 1985)
– For sacroiliac support the transverse abdominis provides anterior compressive force through its attachment to the ilia (Snijders et al 1995; Richardson et al 2002)
• Multifidus
– Superficial fibres cross up to 5 segments whereas deep fibres cross only 2 segments
– Deep fibres are ideally placed to control shear and torsion (Panjabi et al 1989; Bogduk 1997) through
• the generation of compression (Wilke et al 1995)
• control of anterior rotation
• tensioning the thoracolumbar fascia (Gracovetsky et al 1977)
• Other intrinsic muscles
– posterior fibres of iliopsoas, medial fibres of quadratus lumborum, lumbar portions of
longissimus and iliocostalis
– Interspinalis and intertransversarii have a high density of muscle spindles suggesting a major
proprioceptive rather than motor function (Nitz & Peck 1986)
– Internal oblique has fibre orientation similar to transverse abdominis in the pelvis and may
contribute to force closure of the SIJ (Snijders et al 1995)
Sensors
• Muscle spindles contain nuclear bag and nuclear chain fibres which are sensitive to muscle length
and/or velocity of lengthening
• The control component of these intrafusal muscle fibres allows the CNS to adapt the sensitivity of
the spindle elements as well as to adapt the spindle to changes in muscle length
• Alpha-gamma co-activation is essential for the perception of movement (Gandevia & McCloskey
1976)
• Golgi-tendon organs provide inhibitory input to alpha motoneurons and attach to small populations
of muscle fibres which make them sensitive to small forces thereby providing discrete detection of
tension in different parts of the muscle (Houk & Simon 1967)
• Joint receptors are encapsulated receptors (Ruffini and Pacinian corpuscles) which tend to fire at
end of range when the joint is stretched (Nade et al 1987)
• Ligamentous receptors and mechanoreceptors in the annulus of the disc whose electrical
stimulation modulates activity in muscles of the spine including the multifidus (Indahl et al 1995)
• Tactile receptors in the skin include Pacinian corpuscles, Meissner corpuscles, Merkel cells and
Ruffini endings
• Vestibular apparatus includes the saccule and utricle of the otoliths which detect the position of
the head with respect to gravity, and the semicircular canals which detect acceleration of the head
around three orthogonal axes
• ?gravity receptor in the region of the kidneys? (Mittelstaedt 1996)
• Visual and auditory
Controller
• Corticospinal input to the trunk muscles, course the spinal cord bilaterally or send collaterals to
both sides (Kuypers 1981, Mori et al 1995)
• More significant control of the trunk muscles by the brainstem such as the reticulospinal and
vestibulospinal tracts which is consistent with the relatively small representation of the trunk muscles on the motor and sensory homunculii (Kuypers 1981)
• The CNS has three primary strategies for the control of movement
– Feedforward and Open-loop strategies where the outcome of movement is predictable (e.g. ballistic and repetitive movements and predictable challenges to trunk control such as limb movements {Taub & Berman 1968})
– Closed-loop where sensory feedback is required in novel unpredictable tasks (Schmidt & Lee 1999)
– Underlying level of tonic activity to increase muscle tension and hence stiffness acting as a line of defence against unexpected perturbations (Johansson et al 1991) using components of both feedforward and feedback strategies
• The CNS draws on the architectural characteristics of the trunk muscles to meet the concurrent demands of mobility and stability (Hodges 2005)
• Motor programme theory
– Involves memory based mechanism whereby a generalized programme is stored as an abstract representation of a group of movements that are retrieved when movement is performed (Schmidt & Lee 1999)
– The CNS store invariant features of movement such as sequence, timing, duration, relative force
– However, Bernstein (1967) that too many degrees of freedom needed to be controlled concurrently for even the simplest movement
• Dynamic Pattern Theory (Kelso 1984)
– No central representation of all components of movement, but instead co-ordination is determined by environmental invariants and limb dynamics based on the principle of non-linear dynamics
Lumbo-pelvic stability
• Prediction based on an ‘internal system of body dynamics’ which is an abstract construct built up
over a lifetime of movement experience and holds information about the interaction of internal and
external forces (Gurfinkel 1994)
• Parallel processing model for posture and movement (Massion 1992)
• The temporal and spatial features of the superficial muscles are linked to the direction of forces
acting on the spine (Aruin & Latash 1995; Hodges & Richardson 1997) and are therefore linked to
orientation control
• In contrast the deep muscles such as transverse abdominis and multifidus contract independent of
the direction of movement and reactive forces (Moseley et al 2000) and therefore are link to
intersegmental stability
• Attention-demanding tasks produces a latency in the superficial muscle activation whereas no
such latency was found in the deeper muscles (TrA Hodges & Richardson 1999, deep fibres of
multifidus Moseley et al 2001)
• The CNS predicts the amplitude and adjusts the feedforward response accordingly
Closed Loop control of the trunk
• Intended movement is compared against feedback regarding the status of the body and its
relationship to the environment. If feedback differs from the intended movement an error signal is
generated to correct the movement performance (Schmidt & Lee 1999)
• May operate at a basic reflex level using mono-synaptic stretch reflexes for example short latency
reflexes when catching an unexpected mass (Moseley et al 2001) which act ‘en masse’ with no
differentiation between deep and superficial component (Moseley et al 2001). There appears to be
some distant integration as loading or unloading of the arms can produce this trunk response and
can be enhanced with predictability of the loading
• Electrical stimulation of the SIJ in cats (Indahl et al 1999) and annulus fibrosis in pigs (Holm et al
2000) elicits short latency responses on both sides and over multiple segments.
Motor Control - Long loop reflexes
• Long latency involving processing at the cortical level and are thought to provide a greater role in
error correction (Schmidt & Lee 1999). For instance when a supporting surface is suddenly removed, a complex interplay of several body parts, including responses of trunk muscles is activated in addition to using a hip or ankle strategy (Horak & Nashner 1986)
• Used for long duration tasks that require accuracy whereby sensory information is used by the
CNS to continually modulate movement performance in a goal directed manner
Control of muscle stiffness
• Muscle stiffness related to feedforward and feedback mechanisms provides joint support through
the modulation of muscle tone and contributes to control even before the shortest reflex response
could be initiated (Bergmark 1989; Johansson et al 1991)
Effect of Pain and Injury on Motor Control
Changes in open-loop control mechanisms
• People in remission for their chronic low back pain demonstrated delayed onset of TrA with arm
and leg movements in all directions (Hodges & Richardson 1996, 1998)
• Data suggest that the responses are a result of inappropriate motor planning rather then changes
in excitability or transmission of the command in the CNS (Hodges 2001)
Changes in the closed loop control mechanism
• Changes in all elements of the closed loop system have been reported. It is frequently difficult to
determine whether the problem lies with inaccurate feedback from the periphery, inability to
interpret feedback or inability to initiate an appropriate command (Hodges 2005)
• Sensory deficit can be detected by measuring the acuity or smallest perceptible stimulation and to
accurately copy a movement and return to a position after it has been demonstrated by the same
or opposite limb
– Impaired acuity to spinal motion in low back pain (Taimela et al 1999) and inability to reposition with low back pain (Gill & Callaghan 1998) has been demonstrated
– This can lead to delayed reflex response as a result of increased time to reach a threshold response, faulty error detection and the development of a faulty ‘internal model of body dynamics’
– Muscle spindle sensitivity may also be altered by pain (Pederson et al 1997)
• Afferent inputs may be misinterpreted in cases of hyperalgesia and allodynia
• Changes in muscle activity may affect acuity (Gandevia et al 1992) and deep muscles attach to
joint whereby contractions may affect sensation
• Fatigue may also affect sensory acuity (Carpenter et al 1998)
• Considerable debate as to the presence of augmented paraspinal muscle activity with some
reporting increased (Wolf & Basmajian 1977, Arena et al 1989), reduced (Sihvonen et al 1997),
asymmetrical (Cram & Steger 1983) and change in activity (Collins et al 1982)
• Consistent finding of sustained erector spinae activity at end of range flexion when they would
normally be inactive due to the ‘flexion-relaxation’ phenomenon (Shirado et al 1995, Zedka et al
1999, Kaigle et al 1998) and reduced relaxation during gait (Arendt-Nielsen et al 1996)
• Delayed reduction in erector spinae muscles when the trunk is unloaded (Radebold et al 2000)
• Further impairments seen are
– balance when standing on one leg (Luoto et al 1998) or two legs (Byl & Sinnott 1991) or
sitting (Radebold et al 2001)
– increased risk of developing low back pain and recurrence has been shown in people with
poor standing balance (Takala & Viikari-Juntura 2000)
– people with low back pain having slower reaction times (Luoto et al 1995) which has been
associated with predisposition to injuries in many sports (Taimela & Kujala 1992)
– in multifidus fibre composition (Rantanen et al 1983), increased fatigability (Roy et al 1989,
Biederman et al 1991) and reduced cross sectional area, identified as early as 24hours after
the onset of acute unilateral low back pain (Hides et al 1994)
• Therefore, the deep and superficial trunk muscles are affected in an opposite manner in the
presence of pain
Effect of Pain and Injury on Motor Control
Motor control – cause or effect of dysfunction?
• Changes of motor control can be as a result of pain or incompetent motor strategies lead to
inefficient spinal control, thus microtrauma, nociceptive stimulation and pain (Janda 1978, Farfan
1973, Cholewicki et al 1997)
• Studies have demonstrated changes in motoneuron excitability (Matre et al 1998), decreased
cortical excitability (Valeriani et al 1999) and changes in sensitivity in muscle spindles (Pedersen et
al 1997),
• However, changes in motor planning and not simple inhibition or transmission delays have been
suggested (Hodges 2001) and is consistent with the finding that pain changes the activity in the
regions of the brain involved with motor planning (Derbyshire et al 1997) due to the attention
demanding nature of pain or stress associated with pain thereby affecting limited resource
processing capacity (Luoto et al 1999), however change in control of rapid arm movement tasks
was not affected by attention-demanding nor stressful tasks (Moseley et al 2001)
• Fear can replicate some features of the changes demonstrated in clinical and experimental pain
(Moseley et al 2001)
Pain Adaptation Model
• Movement velocity and amplitude is reduced in the presence of pain (Lund et al 1991)
• In the limb and jaw this is represented by reduced agonist and increased antagonist activity
(Svensson et al 1995), in the trunk this may be represented by excessive splinting of the superficial
muscles as predicted by Panjabi (1992)
• Alternatively pain may affect motor control indirectly through its influence on proprioception,
whereby in chronic pain non-nociceptive mechanoceptors contribute to afferent input on second
order nociceptors thereby providing inaccurate feedback affecting feedforward responses due to a
faulty ‘internal model of body dynamics’
Motor control – task conflict
• Task conflict between respiration and pelvic floor
• Respiratory and genitourinary problems are common in people with low back pain (Finkelstein
2002)
Respiration
• If respiration is increased involuntarily (as in hypercapnia) transverse abdominis is recruited at
lower minute ventilations than the other abdominal muscles (Hodges et al 1997)
• During repetitive arm movements, tonic activity in the diaphragm and TrA is maintained but varies
with respiratory demand (Hodges & Gandevia 2000)
Continence
• Increased intra-abdominal pressure form muscle contraction exerts pressure on the pelvic floor
such as coughing (Deindl et al 1993) and lifting (Hemborg et al 1985) and during repetitive
movements of the arm (Hodges et al 2002)
• Volunatry activation of the pelvic floor is associated with involuntary activation of TrA (Sapsford et
al 2001, Critchley 2002) and conversely TrA activity is associated with pelvic floor muscle
recruitment (Sapsford & Hodges 2001)
Trunk may be recruited as a reference frame as in dancing (Mouchnino et al 1993)
Adjacent areas may influence trunk recruitment
• Thorax
• Hips – reduced motion seen in people with low back pain (Ellison et al 1990)
Motor Control and Pain
• Under normal circumstances pain is the result of stimuli that either threaten or result
in injury (Willis 1989)
• Sensory – discriminative and motivational-affective pathways operate in parallel (Melzack & Casey 1968)
• Even the memory of pain can condition behaviour “fear-avoidance’ which may serve
a purpose to avoid further injury during acute pain but can be deleterious in chronic conditions.
• The majority of investigations into pain have been done on cutaneous receptors which appear to act quite differently to deep receptors such as those from joints and muscle (Galea 2005)
• Further difficulties arise in stimulus control as well as interpretation of motor behaviour due to the variety of conditions and the heterogeneity of the response
• Muscle pain induced by an injection of a 5% solution of hypertonic saline (Kellgren 1938) appears to have similar behvioural manifestations as those seen clinically (Arendt-Nielsen et al 1996)
• Cutaneous receptors terminate in lamina I & II of the dorsal horn, whereas deep receptors from muscle and joints terminate in lamina I and/or IV and V (Mense 1986, Hoheisel et al 1989)
• Nociceptive specific neurones (NS) are found predominantly in lamina I and the outer part of lamina II, whereas wide-dynamic range (WDR) neurones are more concentrated in lamina V
Pain
• Convergence – Projection theory (Ruch 1946) is based on the multiplicity of overlap of receptive
fields in the dorsal horn which may explain the referral of deep pain to cutaneous regions and the
diffuse nature of deep pain sensations
• The more medially located the cell is in the dorsal horn, the more distal is the site of the deep
receptive field (Yu & Mense 1990)
• Nociceptors are found throughout skeletal muscle but are most abundantly found in the tendons,
fascia and aponeuroses (Stacey 1969)
• Lesions of muscles can lead to release of endogenous sensitising substances such as kinins and
prostaglandins which may cause oedema increasing pressure on the venous system resulting in
ischaemia which promotes the release of Prostaglandin E2 and Bradykinin. Ischaemia may lead to
failure of the calcium pump and local contracture further impairing circulation which is a theory
behind the development of trigger points.
• Sensitization of muscle tissue can lead to enhanced excitability of dorsal horn neurones which can
cause central sensitization.
• The ‘vicious cycle’ terminology has been given to the painful lesion stimulating gamma motoneurons
which leads to enhanced muscle spindle sensitivity. Enhanced spindle activity activates the alpha
motoneuron through a mono-synaptic reflex causing hypertonus of the affected muscle (Johansson
& Sojka 1991)
• Joint pain may arise from stimulation of nociceptors in the joint capsule, ligaments, bone, periosteum, articular fat pads and around blood vessels but not in the joint cartilage (Wyke 1981)
• Increased background activity as well as increased activity to noxious and non-noxious movement in
A beta, A delta and C fibres occur as a result of inflammation. ‘Silent nociceptors’ begin to respond in
this scenario (Schaible & Schmidt 1988)
Pain & the Brain
• The thalamus represents the final link in the transmission of impulses to the cerebral cortex,
processing almost all sensory and motor information prior to transfer to cortical areas
• Spinothalamic afferents mediate sensory discriminative information and terminate in VPL nucleus
and medial thalamic nuclei, whilst receiving extra information from the globus pallidus, reticular
formation and the cerebellum (Galea 2002). Diffuse cortical projections from thalamic intralaminar
nuclei have been considered as part of a non-specific arousal system.
• The basal ganglia are associated with planned action (Brooks et al 1993) and movement
(Colebatch et al 1991). Their connections through the thalamus, prefrontal cortex, supplementary
motor cortex, motor cortex and anterior cingulate cortex (Cote & Crutcher 1991) form a circuit
associated with motor preparation or response selection.
• A number of brains stem structures, including peri-aqueductal grey matter (Bernard & Bandler
1998) and the reticular formation have extensive connections with all levels of the nervous system
and are involved with nociceptive, autonomic and motor systems
• Multiple cortical areas are activated by pain. These include the primary somatosensory cortex
(Bushnell et al 1999), the secondary somatosensory cortex, the pre-frontal cortex, anterior
cingulate cortex (Talbot et al 1991), and supplementary motor area (Coghill et al 1994)
• Parietal areas are mainly concerned with sensory-discriminative aspects whereas frontal-limbic
connections subserve the affective dimension of pain experience (Galea 2005)
• There are corticospinal projections to all lamina of the dorsal horn from mainly post-central cortical
areas, whereas pre-central cortical areas project to lamina VII and VIII, with neurones from the
motor cortex projecting to lamina IX (Galea & Darian-Smith 1997, Maier et al 1997)
• The majority of these descending projections terminate on interneurons which are part of the
spinal circuit which means that the ‘sate’ of the interneuron is dependent upon the combined
descending input, afferent input, and spinal interactions for the upcoming motor task
• Corticospinal projections to the superficial lamina can modulate NS neurons (Cheema et al 1984)
• Posterior parietal cortex has connections with primary somatosensory cortex and other polymodal
associated areas, including the limbic system (Cavada & Goldman-Rakic) and is part of a general
attentional system.
• The cingulate cortex is involved in the affective and motor behaviours (Devinsky et al 1995). This
region contains neurons which fire in anticipation of pain and therefore could be involved in
avoidance behaviour (Koyama et al 1998)
• The insula receives converging information from all 5 sensory modalities and has extensive
connections with the limbic system and spinal cord (Galea 2002)
Reflexes
• Stretch reflex
– Ia fibre monosynaptic excitatory reflex to agonist and disynaptic inhibitory reflex to antagonist. Gamma motoneurons regulate the sensitivity of the muscle spindles (annulospiral endings),
– May be facilitated during painful stimuli by sensitising Ia muscle spindle afferents (Pedersen et al 1997; Wang et al 2000), however inhibition has also been demonstrated by Mense & Skeppar (1991)
• Tension reflex
– Golgi tendon organ group II fibres make disynaptic inhibitory connections with agonist and facilitatory with antagonist
• Group II reflexes
– Flower spray endings of nuclear bag and nuclear chain fibres detect changes in length and the rate of change in length respectively
– Make disynaptic connections onto motoneurons
– Excitatory reflexes are directly mainly at the flexors and inhibitory connections to the extensors
• Flexor Reflex
– Withdrawal reflex to noxious stimuli (Sherrington 1910), whereas Lundberg et al (1987) promoted the flexor reflex during normal movement
• Withdrawal Reflex
– Noxious stimulation of joint or muscle tissue evokes a limb flexion reflex (Mense 1986)
Pain & the Brain
Muscle hyperactivity and chronic pain
• Facilitation of gamma reflex through group III and IV mechanoceptors and nociceptors in painful
muscles was proposed by Johannson & Sojka (1991)
• This facilitation in turn promotes excitation of group I and II afferents which influence the alphamotoneurons causing further contraction, possible ischaemia and release inflammatory
metabolites such as Bradykinin and Prostaglandin E2
• Increased concentration of inflammatory metabolites from muscle contractions, including lactic
acid, potassium chloride (Jovanovic et al 1990), arachidonic acid (Djupsjobacka et al 1994),
bradykinin (Pedersen et al 1997) and 5-HT (Djupsjobacka et al 1995) results in increased gamma
fibre activation and sensitivity of muscle spindles leading to increased stiffness.
• Experimentally induced pain has been shown to be associated with changes in the fusimotor
system (Thunberg et al 2002)
• Neck and paraspinal muscles are rich in muscle spindles (Richmond & Abrahams 1975, Amonoo-
Kuofi 1982, Boyd-Clark et al 2002). These spindles, many of which lack bag fibres, are controlled
by the static fusimotor system and are mainly concerned with postural muscle activity (Price & Dutia 1989) and therefore disturbances in these fusimotor fibres are likely to cause disturbances in motor co-ordination and proprioception
• Stimulation of fusimotor activity through the application of vibration to the neck muscles has been
shown to cause disturbances in balance and motor activity (Lund 1980)
Muscle hypoactivity and chronic pain
• Clinical observation also suggest that there can be inhibition of muscle activity
• An alternate model, ’the pain adaptation model’ proposes that muscle dysfunction is a normal
protective adaptation and is not one of the causes of pain (Lund et al 1993)
• Investigations suggest inhibition of the agonist and facilitation of the antagonist during walking
thereby reducing movement amplitude and stride time during experimentally induced leg muscle
pain (Graven-Nielsen et al 1997; Madeleine et al 1999)
• Injection of hypertonic saline into one side of the erector spinae has been shown to reduce the
velocity and range of voluntary trunk movements (Zedka et al 1999), where the concomitant
reduction in EMG amplitude persisted even when subjects pushed through the discomfort
• Patients with chronic low back pain fatigue faster (Kankaanpaa et al 1998) and have poorer
balance performance and delayed postural response times (Radebold et al 2000, 2001)
• In normal subjects, fatigue can result in altered anticipatory adjustments (Allison & Henry 2002)
Wider effects
• Injection of hypertonic saline into the trapezius muscle during low load repetitive work task not
only resulted in reduced activity in the injected muscle but additionally altered rhythm, a tendency
to increase the amplitude of arm movements and a prolongation of the role of the non-affected
arm in the task (Madelaine et al 1999)
• Injection of hypertonic saline into the soleus muscle altered gait and hence rhythm of other
muscles in the synergy (Martin & Arendt-Nielsen 2000)
Models of Motor Control
• It is now realised that the distinctions made in the hierarchical model (upper motor neurones for
purposeful action and lower motoneurons for reflexes) and the simple stretch reflex are insufficient
to explain simple models of motor control as these distinctions become more and more blurred.
For example every voluntary movement has a concomitant reflexogenic postural adjustment that
occurs subconsciously
• Central pattern generators (CPG) are derived from motor programs where a sequence of
commands have been pre-programmed.
• Examples of CPGs are rhythmical movements such as walking and respiration which are motor
patterns that receive sensory feedback thereby making it a closed loop system
• Open loop system require feedforward mechanisms which deliver information to other parts of the
system to ‘prepare’ it for upcoming motor commands such as postural adjustments prior to limb
movements
• Bernstein (1967) perspective described movement constrained to synergies or coordinative
structures (ie functional units of joints and muscle constrained to act as one) whereby control is
not exerted over muscles or sensory receptors (reflex model) nor muscle activation patterns
(hierarchical model) but rather over abstract relations between kinematic variables and the
accomplishment of task goals
• The Dynamic Action Theory (an elaboration on systems theories) proposes that movement
emerges naturally out of complex interactions among many interconnected elements (physical,
environmental and neural) without specific commands or motor programs in the CNS
• At the core of dynamic action theory is that human behaviour is governed by a generic process of
self-organisation; functionally linked rather than mechanically linked (Kelso & Tuller 1984), task
dependent and constrained as units of action specified in dynamic terms rather than kinematic or
muscular variables and determined by features of the action such as movement trajectory
(Saltzman & Kelso 1987)
Nervous System Plasticity
• Cortical plasticity has been most dramatically demonstrated by Merzenich and colleagues (1983,
1984) whereby digital representation of the hand was remapped in the somatosensory cortex
following deafferentation and digital amputation in monkeys.
• In humans this re-organisation is not haphazard but context specific whereby the brain can be
viewed as a dynamic entity (Kelso 1995)
• Rapid repetitive highly stereotypical arm movements in monkeys were shown to degrade sensory
representations of sensory information guiding fine motor hand movements leading to dystonia
and subsequent lack of use of the hand (Byl et al 1996)
Dynamical View of Pain and Motor Control
– Groups of cells rather than single cells are the main units of activity in the nervous system
– Functional synergies of muscles rather than single muscles are then main units of motor control and
coordination of action
– There is distributed control of these systems
Timing relationships between muscles appears to be the critical restraint rather than amplitude.
Chronic Pain and Motor Control
• Pain is essentially motoric, whereby the basic biological function of pain is to escape or prevent
danger and promote survival (Melzack 1996, Wall & Melzack 1999)
• Abnormal movement patterns can be compensatory to pain, causative of pain or neither
• What defines chronic pain still has not been established – mechanisms/aetiology, duration,
physiology
• The complexities between chronic pain and movement concern the lack of understanding of the
physiological complexities of chronic pain and the potent effects of cognitive and emotional factors
• Allodynia has particular implications as non-noxious movements are now deemed painful
suggesting that even small deficits in motor control may be sufficient to maintain a nociceptive
input
• Peripheral nerve injury can induce more robust peripheral hyperalgesia whereby small regenerating sprouts who don’t reach their target organ, turn back on themselves creating a small tangle of nerve fibres which can become ectopic impulse generators (Devor & Seltzer 1999, Wall & Gutnick 1974, Xie et al 1993) are sensitive to tension & compression, chemical substances as a result of anoxia and inflammation and noradrenaline (Janig et al 1996)
• The dorsal root ganglia (DRG) may also become a source of ectopic generators whose afferent
input can sensitise the dorsal horn of the spinal cord, and whose efferent input can create neurogenic inflammation which may result in secondary hyperalgesia
• Unlike the rest of the peripheral nervous system, the dorsal root ganglion is particularly permeable
to vascular circuits, thereby making circulating chemicals and humoral factors particularly accessible. This has important ramification to people in chronic pain who often suffer high levels of stress with the associated release of catecholamines
• Finally, the DRG is situated close the intervertebral disc where damage to the disc can release
inflammatory cytokines which may permeate into the DRG
Central Mechanisms of Chronic Pain
• Reduction of normal inhibitory inputs (‘disinhibition’) caused by removal of supraspinal projections
to the WDR neuron can cause sensitization (Doubell et al 1999) suggesting that pain sensitisation
can occur in the absence of peripheral mechanisms. Furthermore, increased descending facilitation of interneurons can also result in pain.
• These changes are called central sensitization and manifest clinically as secondary hyperalgesia
and allodynia which is produced from elevated resting membrane potential, reduced activation
thresholds of the second order nociceptor, altered gene expression and sprouting at the receptive
synapses (Dubner & Ruda 1992)
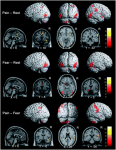
• Cortical structures are known to undergo structural reorganisation in patients in whom no peripheral neuropathy is demonstrable. Herta Flor and colleagues (1997) investigated the extent of somatosensory activation in people with chronic low back pain by showing noxious stimulation to increase somatosensory activity as a function of the duration of their pain. However, other studies demonstrate variable results
• The most consistent finding across studies is the activation of the anterior cingulate cortex (ACC)
(Peyron et al 2000) which is thought to be important in anticipation of pain (Sawamoto et al 2000)
and anxiety (Kimbrell et al 1999). The ACC appears to be constantly active in chronic pain (Hsieh
et al 1995)
• It seems that the ACC is part of the motor function whereby the brain makes sense of the
nociceptive input according to what it should do about it (Wall 1999)
• ACC is important in establishing an emotional valence to pain and co-ordinating the perception of
bodily threat with other areas in selecting and planning the appropriate behavioural/motor
response strategy (Price 2000).
• The ACC projects directly to motor and supplementary motor areas (Price 2000), and receives
direct inputs from reticular formation neurons having a relevance in escape behaviour and fear
response (Willis 1985)
Anterior Cingulate Cortex (ACC)
• Factors that are thought to impart threat or emotional valence to pain are thought to have greater
impact on motor control than factors that impart stimulus intensity or sensory qualities to pain. As
cognitive and emotional factors contribute more to pain, so too the impact of pain on motor control
increases (Price 2000)
• The majority of pre-synaptic projections from the dorsal horn are from WDR neurons is significant
in that sensitisation of the dorsal horn almost exclusively involves WDR neurones promoting escape type responses
Implication of neural changes
•The neural adaptation associated with many chronic pain situations can be demonstrated as normal
adaptation in abnormal circumstances
•Increased nociceptive sensitivity leaves a smaller kinetic margin for error
•Tolerance to non-nociceptive input is reduced due to WDR sensitivity, therefore creating a fine line of
just doing enough to promote strength, endurance and control without doing too much resulting in a so
called ‘flare up’ typically associated with severe pain sometimes with paraesthesia and nausea
•Cortically mediated modulation of nociception may be sufficient to increase and maintain pain where
perceived threat and sense of vulnerability are central to this effect
•Structural reorganisation of the CNS can result in inaccurate transmission of proprioceptive
information, which may disrupt internal spatial representation of the body, thereby further compromising motor control
Motor control in reducing lost game time in Australian Rules Footballers
A motor programme delivered to elite Afl players was effective in retaining muscle size and function of the multifidus and transverse abdominis whilst reducing the number of games missed during the season (Hides et al 2012, Med Sc Sp Ex, 44, 6, 1141-1149)
see Pain in the Brain Imaging elsewhere on this site
18 August 2012
 What We do
What We do What We Treat
What We Treat