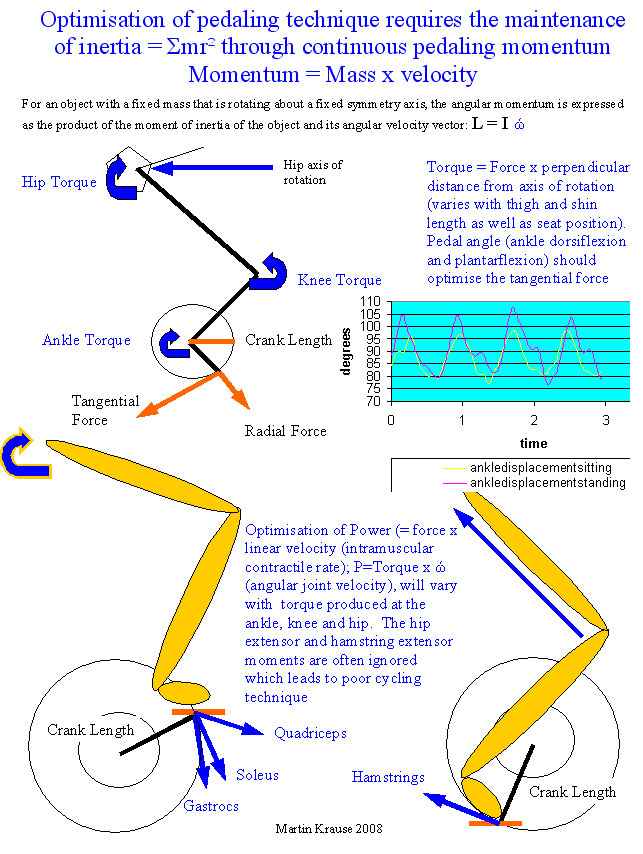
Injury prevention, education, and performance through the analysis of cycling technique
by Martin Krause
Muscle coordination patterns for efficient cycling is achieved through coordination of muscles at the same joint, across several joints through sequential peak activation patterns from knee to hip to ankle, and reliance on multiple muscles for large joint torques. Variation of activation patterns at top and bottom dead center are also suggestive of important coordination patterns between left and right legs (Blake et al 2012, Med Sc Ex Sp, 44, 5, 926-938). At Back in Business Physiotherapy we specialize in cycling optimisation and comfort. Whether you uncomfortable on your bike, or have a specific injury, and/or would just like to improve your performance, we will do a thorough examination. This examination would look at posture, bony alignment, muscle length, strength and synergistic timing, as well as aberrations of movement. Our approach uses releases of myofascial trains, joint mobilisations and manipulations, muscle energy techniques, acupuncture dry needling, taping as well specific exercise regimes. This multidimensional and multimodal approach has been developed since my involvement with cycling since the 1970's and my association with The Rennbahn Klinik and Magglingen in the 1980's where we treated and screened professional Tour de France Cyclists, and Elite Amateur Swiss national team cyclists in preparation for the Barcelona Olympics in 1992 as well as the Sydney Olympics in 2000. Furthermore, cycling has been shown to have potent effects in the treatment of chronic low back pain.
Index
Kinematic comparative analysis of pedaling in seated versus standing cycling
by Martin Krause 2004
Task Demand
Cycling requires efficient pedaling action to maintain adequate power output to win a race. Importantly, an endurance sport such as cycling requires a pedaling action, which provide sufficient cadence to maintain a steady state of efficiency. In particular race advantage over competitors can be achieved through efficient hill climbing. Anecdotal evidence suggests that cyclists prefer to stay seated as long as is possible before rising out of the saddle. However, acceleration during hill climbing is frequently achieved through standing off the saddle. This acceleration appears to be achieved through the use of body weight and arm moments as well as increased use of shank and thigh movements. Therefore, increased plantar flexion and hip-knee extension may be expected.
When considering moments of inertia it is probable that efficient cycling technique involves the ability to maintain saddle contact even during hill climbing. Additionally, when considering the sliding filament model of muscle action, it is likely that due to specificity of muscle contractile action it will be the cyclist who can maintain a pedaling action which does not vary considerably from sitting to standing who will be most efficient in hill climbing action. However, power may be a greater consideration than efficiency when standing out of the saddle. Therefore, effective pedal force may also be the result of arm action. Finally, when comparing body position from the seated to standing position, the cyclists center of gravity moves further forward and therefore more likely to be over the center of crank rotation in the standing position.
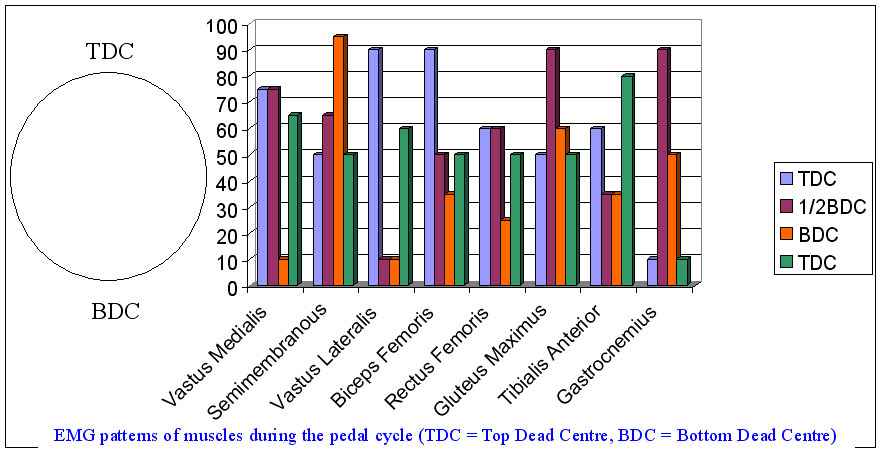
At the bottom of the pedal cycle, during the transition period between the downstroke and upstroke, the ankle plantar flexors and the knee flexors (Semitendonosis and Biceps) showed the greatest EMG activity. Tibialis Anterior (TA) and Rectus Femoris (RF) have demonstrate large EMG activity at TDC, whereas Vastus Lateralis (VL), Vastus Medialis (VM) and Medial Gastroc (MG) have increased EMG activity during the downstroke. Therefore progression of muscular activity during the downstroke are TA and RF at TDC, VM, VL and GM acting synchronously in the downstroke, and followed by GM, then Soleus, and then LG, MG, ST and BF synchronously (Blake et al 2012). Peak muscle activity occuring sequentially from the knee to hip to ankle with multiple muscles producing large joint torques.
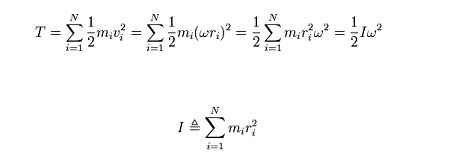
Introduction
Efficient pedaling requires the effective transfer of energy from the lower limbs into the pedal. Consequently, this energy is transferred into the crankshaft, which drives the chain that accelerates the bike. Importantly, the effective use of force, through the pedal at the correct point in time of the crank cycle, is required. Anecdotally, this force is thought to be applied perpendicular to the pedal crankshaft during the down stroke. However, the foot is seldom perpendicular to the pedal crankshaft (Cavanagh et al 1986). Additionally, as the cyclist moves from the seated to the standing position, logic would dictate that the pedal – crankshaft angle will change and the range of the down stroke where maximum force is applied will also change. Finally, since the pedaling action is a cyclic activity then the recovery phase (upstroke) of cycling requires efficient use of energy through the inertial forces of the pedaling action (Cavanagh et al 1986). However, in standing these inertial forces change as the joint torque changes. Therefore, energy can be lost through incorrect pedaling angle resulting in reduced effective force.
Literature Review
Investigations examining the kinematic parameters of seated versus standing cycling are sparse. Most studies have focused on cycling in level terrain (Caldwell et al 1999). Why cyclists adopt the standing position remains unresolved (Davison et al 2000, Ryschon et al 1991). However, the inertial properties of cadence and quasi static forces of lower cadence may be the deciding factor as to whether a cyclist determines the need to stand. Lower cadences would appear to be less efficient and therefore would require greater force to be applied through the pedal (since Power = Force x velocity). Indeed, peak resultant pedal force and peak crank torque increased by 200% and 130% respectively in the uphill standing position (Caldwell et al 1999). Stone and Hull (1993) suggested these forces to represent greater than 60% body weight in the standing position. Furthermore, these peak forces occurred later in the crank cycle for the standing position (at 155 degrees for force and at 130 degrees for torque) than in the sitting position (100 degrees for force and 85 degrees for torque) (Caldwell et al 1999) (see Figure 1). Interestingly, the higher peak kinetic values were linked to kinematic changes in pedal orientation and therefore changes to force vector direction throughout the crank cycle (Caldwell et al 1999).
Investigation into kinematic parameter variations between various seat heights suggest that the peak knee extension increases from 65 to 125 degrees in the low seat to 25 to 105 degrees in the high seat configuration. Additionally, ankle angle varied from 5 dorsiflexion (DF) to 25PF degrees in the low seat condition to 12 plantar flexion (PF) to 20PF degrees in the high seat condition. Peak knee extension and flexion occurred at 170 degrees and 350 degrees of the crank cycle respectively (Gregor et al 1996). This suggests that at least in the varied seated position the muscles length tension relationships would change, thereby affecting force capabilities of the muscle by placing them in a different area of their length-tension and force-velocity curves (Gregor et al 1996). Extrapolation of these results to the standing position, suggest similar implications for the length-tension and force-velocity relationship for the production of power during uphill climbing. However, hip and knee angular kinematics have opposing effects and therefore result in minimal muscle length change in the hamstring muscle (Rugg et al 1987). A similar pattern was seen in the gastrocnemius muscle. In contrast, changes in the length-tension relationships of the mono-articular muscle such as soleus may be expected as it does not cross both the knee and ankle. The magnitude of initial lengthening was about the same in the various seated positions, however the magnitude of shortening increased dramatically from about 1cm to approximately 2.5cm (Rugg et al 1987). Therefore, an expected kinematic outcome for the standing position is increased plantar flexion. This should result in an increased rate of shortening of the soleus muscle from it's initially lengthened position.
Impulse occurs earlier in the crank cycle than does the development of the effective pedal force (figures 1 and 2). Therefore, an increased rate of muscle shortening to accommodate the application of muscle force over an increased range of motion may be seen through altered velocity and acceleration at the ankle joint. Paradoxically, the soleus muscle is considered to be an endurance muscle with slow twitch properties suggesting lesser abilities to shorten quickly (Soderberg 1992). Furthermore, muscles with a large proportion of it's myofilaments in series are considered to have a greater propensity to shorten with greater velocity over a greater range of motion. However, the soleus muscle has a large proportion of it's myobrils in parallel (Jones et al 1990). Additionally, the soleus muscle has some of the largest angles of pennation (up to 60 degrees) seen in the human body (Soderberg 1992). Large angles of pennation reduce effective force transmission (Lieber 1992). Consequently, the implications of altered loading of such an essentially slow twitch endurance muscle are either changes in the myofibril architecture with training, a reduction in cadence in the standing position, altered timing in the application of torque or synergistic muscle load sharing (Caldwell & Li 1998).
In order to examine changes in joint kinematic parameters we maintained a constant pedaling frequency. Additionally, the effect of load sharing between the soleus and gastrocnemius muscle were examined by looking at the kinematic parameters at the knee and ankle to see if they would result in little change in the length of the gastrocnemius muscle, as is suggested by the literature (Gregor et al 1996). Finally, examination of the ankle joint angle (see figures 3 and 4) during the crank cycle should determine variations to the timing in the application of effective force and therefore torque during the crank cycle, described by Cavanagh et al (1986).
Fig 1: pedal impulse: Since Impulse = force x time, it can be seen from the light shaded area that 50% of the propulsive impulse is delivered between 60° and 120° after TDC (top dead center)
Fig 2: pedal effective force: the light shaded area represents 'unused force' whereas the dark shaded area represents negative effective forces. The objective of the rider is to reduce the size of the unused force and eliminate the negative effective forces without decreasing the size of the propulsive impulse
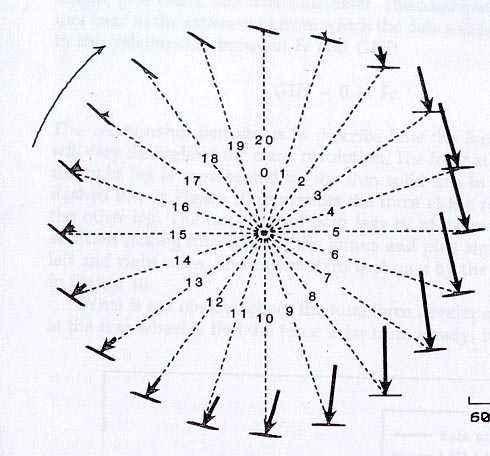
Fig 3: pedal angle and timing of torque: The orientation of the pedal and the resultant force vector are shown at 20 positions of the crank cycle. It is interesting to note the orientation of the force vector during the first half of the revolution and the absence of pull up forces in the second half.
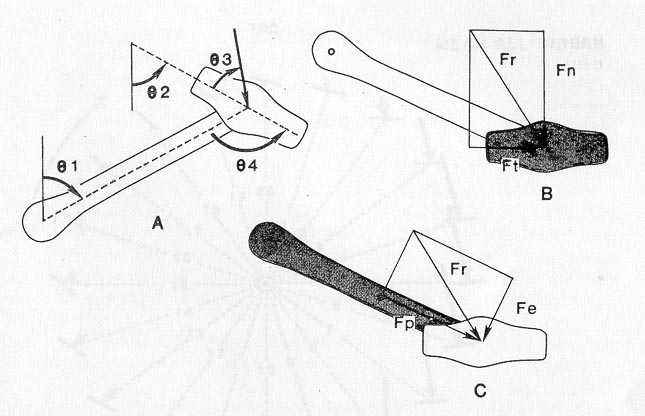
Fig 4: pedal angle: A shows the angle conventions used for the calculation of force. B and C show the resultant force. Fe is called the effective force because it represents that component of force that is effective in generating a force at the rear wheel.
(Figures 1-4 from Cavanagh et al 1986, pp103-109)
Methods
The subject was a healthy 37 years old male amateur cyclist. He was asked to cycle at 60 revolutions per minute in the sitting and standing positions using a bike ergonometer. He had markers placed on the lateral aspect of the shoe at the region of the 5th metatarsal, at the lateral maleolus, at the superior head of the fibula, and at the greater trochanter. A digital video camcorder (Canon Pal MV530i) recording at 50 frames per second was used to record the cycling action. Recordings were made during steady state 60 revs per minute cycling.
The digital video was then captured on the computer using the Ariel Performance Analysis System (APAS) system of kinematic analysis. The captured film sequence was trimmed to remove unwanted sequences, digitized, transformed using the calibration points, and filtered cut off frequency to smooth out errors in digitizing. Once the results were displayed, the data was cut n' pasted into an excel spreadsheet and then graphed for comparative qualitative analysis.
Presentation of results from kinematic analysis
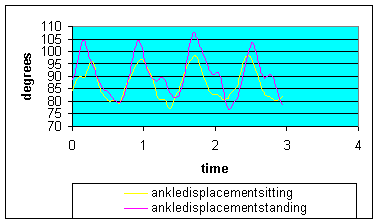
Graph 1: Ankle angular displacement: ankle displacement in standing demonstrated considerably larger (approx 8 degrees) plantar flexion.
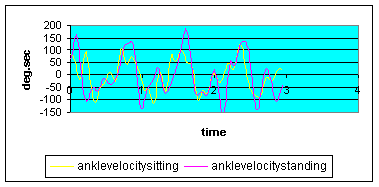
Graph 2: Ankle angular velocity: ankle velocity into plantar flexion during standing was somewhat higher, however the change of direction into dorsiflexion was more pronounced in the standing position
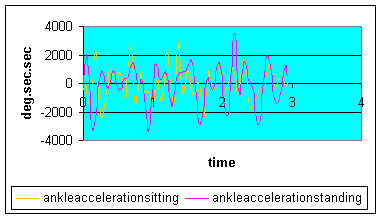
Graph 3: Ankle angular acceleration: ankle acceleration into dorsiflexion (top of graph) appears on average higher than the acceleration in plantarflexion (bottom of graph) during the standing position. This result may be expected as the total displacement of the ankle has increased for the same revolutions per minute. Therefore greater ankle displacement must occur in the same amount of time. Additionally, greater acceleration may be synonymous with greater impulse generation (I=F.t=m.a.t=m.v) and therefore greater momentum. However, the momentum generated will be influenced not by velocity alone but by the greater mass of body weight in the standing position.
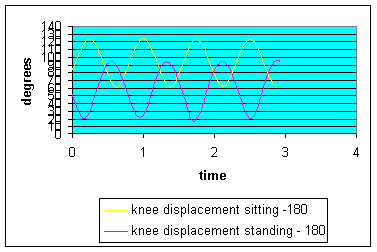
Graph 4: Knee angular displacement: The total range of knee motion did not change significantly from the sitting to standing positions. However, the displacement occurred in a more extended position (bottom of graph) of knee extension in the standing position. Yet the results in the standing position are within the expected range of motion if the seat height is correctly fitted for the rider (see below).
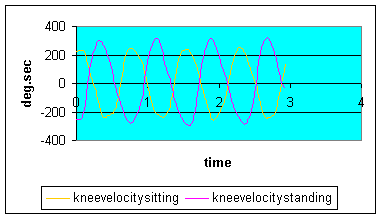
Graph 5: Knee angular velocity: Knee velocity did not vary greatly between sitting and standing as the displacement between the two positions were similar.
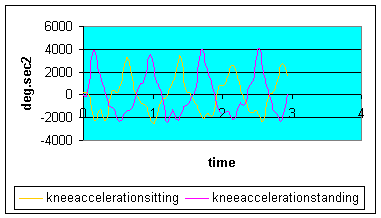
Graph 6: Knee angular acceleration: Knee acceleration did not vary greatly between the sitting and standing positions suggesting that the Impulse generation and momentum were similar at the knee joint in the 2 conditions. These results suggest that a greater impact may be expected on the internal mechanics of the actin-myosin sliding filament in the quadriceps and hamstrings muscle due to their working in a different range of motion. Importantly, since mass is increased through the application of body weight in the standing position, then the velocity of shortening may be expected to decrease.
Discussion of results
Our results suggest that ankle displacement is strongly influenced by changes in body position. The results demonstrate an average range of ankle displacement of 25° (range = 80-104° ) for the standing position and 17° (range = 80-97° ) in the sitting position. Similarly, Caldwell et al (1999) demonstrated 17° of ankle displacement in standing. This compares with 40° 58'+6.01 and 25° 7'+14.12 in sitting and standing respectively found by Shemmell & Neal (1998) and Too (1996). These authors provided normative data suggesting that the range of angular displacement at the ankle in standing was 137° 40'+9.53 to 96° 82'+11.07, with the pattern of angular displacement showing a trend to reach maximum plantar flexion earlier in the pedal cycle in the non-elite group (Shemmel & Neal 1998). This suggests that the optimal pedaling angle during standing is always in some plantarflexion thereby providing an effective force angled downwards and backwards. Therefore, during the recovery phase of the pedal cycle little or no upward pull can occur on the pedal. When considering Figures 1 and 2, this technique should then minimize negative effective forces described by Cavanagh & Sanderson (1986) on the upstroke of the pedal cycle. However, prolonged plantarflexion does suggest an increase of 'unused force' at the bottom of the pedal cycle. Our results vary significantly as our subject demonstrated the same amount of dorsiflexion whether in sitting or in standing. Additionally, this dorsiflexion occurred below the 90° angle suggesting a large amount of eccentric calf muscle activity prior to plantarflexion. In contrast, data from Caldwell et al (1999) demonstrated that regardless of the condition, ankle moment profiles demonstrate exclusively plantarflexor torque throughout the crank cycle, with the highest values after 90° , i.e. in the latter part of the downstroke. Shemmel & Neal (1998) concluded that ankle angles were found to be the best discriminative tool as the ankle plantarflexors contribute more to force production in the standing versus seated position.
Our knee displacement results in standing of 72° and 62° in sitting contrasts significantly with 28° 69'+8.80 and 73° +6.41 (Shemmell & Neal 1998). Caldwell et al (1999) demonstrated knee displacement of 82° 39', with a range from 17° 6' knee extension to 99° 45' knee flexion in the standing condition. Shemmel & Neal (1998) attributed their dramatic change to the forward shift of the cyclist over the pedals during standing. The significant difference between our results and their results may be due to the outdoor 10.5% incline condition, which they used for their analysis. Additionally, they factored the bicycle tilt in the frontal and saggital planes into their calculations. Stone & Hull (1993) suggested a 5° phase lag due to the 8% incline used in their investigation. Finally, Caldwell et al (1999) had their subjects use their own bicycles and were asked them to complete the task as closely as possible to real racing conditions. These latter factors are in contrast with our protocol as we maintained the same incline and cadence; whereas in racing conditions the incline increases and cadence frequently decreases during the uphill phase of the race (82rpm to 65rpm {Caldwell et al 1999}). A notably exception to this rule was an increase in Lance Armstrong's cadence during the 2001 Tour de France on Col Du Madelaine.
Effectiveness of performance
The demonstrated kinematic changes suggest kinetic consequences to work and power production. Caldwell et al (1999) demonstrated an increase in the peak magnitude of the ankle plantarflexor and knee extensor moments during uphill cycling. They suggested that these changes were likely related to the total work done per crank revolution as a consequence of holding power output constant while cadence is decreased.
Our kinematic results tended to suggest that peak plantar flexion occurred earlier in the crank cycle. Yet the results obtained by Caldwell et al (1999) suggest distinct kinetic modifications at the knee and ankle later in the crank cycle in the uphill standing position. Ankle plantarflexion moment increased by 160% and shifted by roughly 45° in the crank cycle. The knee extensor profile also showed a shift towards the late downstroke period, with a bimodal extensor pattern that exhibited a second peak at about 135° crank angle (Caldwell et al 1999). In the uphill standing posture the cyclists body moves forward and upward. The knee extensor phase persists into the first portion of the upstroke, as does the ankle (Caldwell et al 1999). These kinematic changes result in altered pedal orientation to a more toe downward position throughout the crank cycle and modification of the applied pedal vector (Caldwell et al 1998).
Features which contribute positively to performance
In the standing position, the line of action of the force vector is closer to the ankle joint center, but the pedal force is much larger resulting in an ankle joint moment which is increased in the later part of the downstroke. Similarly, in the standing position the force line of action is posterior to the knee in the earlier part of the crank cycle (140° ), but moves in front of the knee later in the downstroke (near 160° ) (Caldwell et al 1999). Unfortunately, we were not able to establish this point because we did not use a body marker in our analysis.
In standing the removal of saddle support results in the greater contribution of gravitational forces. In the later part of the downstroke, the rider can make greater use of the effective force by angling the pedal downwards thus increasing the backward horizontal component of the force. In contrast, in the seated position, the backward horizontal component can only be generated by pulling back on the pedal with a flexor moment (Caldwell et al 1999).
Stone & Hull (1993) demonstrated additional positive power moments through the action of the arms during standing. The arms pulled up and back during the power stroke of the corresponding leg and pushed down and forward during the upstroke. This represents a reversal in direction when compared to the sitting condition. Importantly, the maximum lean of the bicycle corresponds with the maximum power output at about 140° of the crank cycle (i.e. downstroke phase). Because the arms pull up and back at this instant, the normal force would be affected. Therefore, achieving forces greater than body weight is a result of the action of the arms.
Improvements, which could be made to performance
Based on inverse dynamics analysis assumptions have been made that mono-articular muscles contract in the absence of antagonist contraction. However, several authors suggest that mono-articular and bi-articular muscles play different roles in the production of multi-segmental actions such as cycling. In particular, the uphill standing position, which frees the hips from the constraint of the saddle, may play an important role in the development of efficient propulsive forces when cycling uphill. Therefore, conclusions based on inverse dynamics may be less accurate when examining the standing condition.
Changes in the effective pedal force direction due to alterations in body position and therefore joint position suggest alterations in the functioning of bi-articular muscles. The gastrocnemius, rectus femoris and hamstring muscles have been shown to be active during cycling (Li & Caldwell 1998). In particular, the later muscle has been attributed to providing a hip extensor moment through a backward pull on the femur through it's insertion into the pelvis (Lombards paradox). Based on kinematic and kinetic analysis, Fregly & Zajac (1996) concluded that the net ankle and hip extensor joint torques function synergistically to deliver energy (38 joules = 50-82% of energy generated to the limb) to the crank during the downstroke. However, they attributed these net moments to the mono-articular soleus and gluteus muscles. Mono-articular muscle may indeed provide the majority of muscular work at each joint, yet the bi-articualr muscles probably redistribute the energy across adjacent joints (Caldwell et al 1999).
During the upstroke, contrary to popular opinion an ankle-hip flexor synergy is not used. Rather, the ankle extensor torque transfers energy from the crank to the limb in the upstroke (Fregly & Zajac 1996). Suggestions were made that in the presence of increased force through application of body weight in the standing condition that the two energy sources (gravity and ankle extensor torque) could restore the potential energy of the limb in the upstroke (Fregly & Zajac 1996). Therefore, the standing position may provide a more effective lower extremity joint angle that increases effective force production (Too & Landwer 2000). However, it would appear more likely that the effective pedal forces are enhanced by positive torques developed by the arms during uphill standing through the pull upwards on the handlebars during the downstroke and the push forward on the handlebars during the subsequent upstroke (Stone & Hull 1993). The maximum upward pull on the handlebars corresponded with maximum power production at 140° of the crank cycle (Stone & Hull 1993).
Minimizing the joint moment cost function would minimize leg stress and fatigue, thus maximizing propulsive power and cycling performance. However, based on the muscle force-velocity relationship, maximal power is not achieved through high velocities and low loads, nor low velocities and high loads, but rather a moderate load and velocity (Winter et al 1996). The standing condition does not suggest optimization of performance due to the high load and low velocity often employed by cyclists. Suggestions that the bi-articular muscles may be optimizing their length tension relationship in the standing condition don't appear to be supported by the literature. Instead, the bi-articular muscles are thought to act as 'energy straps' transferring energy from one limb segment to another (van Ingen Schauer 1989). Furthermore, the biarticular muscles are considered to control the direction of applied external forces by balancing the relative moments of the adjacent joints they cross (van Ingen Schauer et al 1992). Importantly, bicycle lean and effective force production by the arms (as demonstrated by Stone & Hull 1993) have important implications for the transfer of energy through the body to the pedal.
Why particular features of the task may or may not be beneficial
Anthropometrical differences between cyclists, suggests that the taller cyclist with longer leg lengths (and greater inertial properties) would minimize their kinematic moments through reduced pedaling cadence. Conversely, the shorter cyclist (with shorter leg lengths and lower inertial properties) would minimize their quasi-static moments with a higher pedaling cadence (Too & Landwer 2000). Regardless, a cyclist is more likely to stand up out of the saddle to maintain their inertial properties of cadence through a change in joint moments as well as the addition of body weight to the limbs inertial forces. Additionally, the arms provide considerable positive moments during the power phase of the crank cycle. These moments allow the pedaling force to exceed body weight (Stone & Hull 1993). However, unless there is sufficient neurophysiological control and co-ordination, these increased forces may not contribute to effective pedal force. The bi-articular muscles are thought to not only transfer energy but also control the direction of force production. As stated early the standing position results in a body position almost vertically over the center of rotation of the crank, which may be a means to control the perpendicular distance from the point of force application to the axis of rotation. Hereby, the cyclist may attain greater effective force through the pedal. However, due to the use of the arms greater energy is expended which suggests reduced efficiency as less amount of time can be spent in the standing position due to finite amounts of energy (ATP). Therefore, ideally the cyclists should adjust their seat height in order to obtain minimum kinematic joint moment cost function (Gonzalez & Hull 1989). Tri-athletes tend to have bicycles built whereby their seated position is more forward. They also have the additional benefit of being able to slide the seat forward on the seat post thereby bringing their body closer to the center of the cranks (Price & Done 1997) without the need to stand up out of the saddle. However, in contrast to cyclists, triathletes need to maximize their efficiency in swimming and running as well. The forward seating position may more resemble to kinematics of running thereby opitimising the specificity of training the myo-filaments. Additionally, reduced need to use the arms after the swimming leg may represent better cost efficiency in already fatigued muscles.
Conclusions
Our data demonstrated greatest change in the kinematic parameters at the ankle joint in the sitting versus standing condition. During standing the ankle was in larger and more prolonged plantarflexion. This suggests that the crank-pedal angle and the position in the crank cycle where most of the effective force is applied, varies significantly from the sitting to the standing condition. Although, the knee angle during the crank cycle varies significantly from sitting to standing, the overall displacement was the same in the two conditions.
According to the literature, the standing position is likely to produce greater effective force production through peak moments involving the use of body weight as well as the pull and push of the arms through the handlebars during the downstroke and upstroke respectively. Furthermore, the cyclist tends to bring their body upward and forward over the axis of crank rotation in the standing condition thereby altering joint kinematics. The bi-articular muscles are thought to be involved in the transfer of energy from the upper limbs to the lower limbs. Additionally, the bi-articular muscles are thought to control the direction of effective force application in the standing condition.
It is likely, that the cyclist takes up the standing position to minimize the quasi-static moments which could arise if the power generating capacity of the muscles is being compromised through reduced cadence (P=F.v.). The addition of body weight and arm moments should allow the greater use of impulse (I=F.t.) and therefore momentum (M=m.v.). However, the amount of time that work (W=P.t) can be generated in the standing position is likely to be less due to greater energy expenditure. Therefore, the standing position represents a good compromise by enhancing kinematic parameters to produce greater effective pedal forces, although for a short period of time. When compared with sitting (where peak force is around 90 degrees), most authors agree that the development of peak effective force and power occur later and in the region of 135-140° of the crank cycle during standing. According to the literature, this phase lag is a result of alterations in body position, as well as surface gradient incline and bicycle sideways tilt.
Interestingly, in the seated position, at a cadence of 90rpm, relative hip extension power doesn't increase with increasing power (250W ->800W) requirements. What does increase is the time spent in knee extension and the relative knee flexor power contribution (Elmer et al 2011). Conventional wisdom dictates that the hamstrings, together with the hip extensors and vastus lateralis act as synergistic knee extensors in the seated position. Coyle (1995) suggested that more experienced cyclists distribute the work across more muscular actions to reduce the localized stress. Further considerations of efficiency must include the transfer of power generation between the left and right legs. Indeed, Blake et al (2012) have shown muscle activity variablity between the top and bottom of the cycle at approx 60% VO2Max. As cadence decreases, the time lag between legs has greater potential of increasing. Ideally, cyclists should have a cadence of between 90 and 120 rpm when developing full power. Furthermore, as cadence does increase (90->120rpm) the power contribution of the hip extensors does increase (Elmer et al 2011).
Cycling versus Pilates specific exercise for chronic low back pain
Sixty-four patients with low back pain were randomly assigned to 8 weeks of specific trunk exercise group (SEG), or stationary cycling group (CEG). Self-rated pain, disability, catastrophizing and FAB scores were collected before, immediately after (8 wk), and 6 months after the training program. Clinically meaningful improvements were defined as greater than a 30% reduction from baseline in pain and disability scores. At 8 weeks, disability was significantly lower in the SEG compared with the CEG (d = 0.62, P = 0.018). Pain was reduced from baseline in both the groups after training (P < 0.05), but was lower for the SEG (P < 0.05). FAB scores were reduced in the SEG at 8 weeks, and in the CEG at 6 months. No between-group differences in Fear Avoidance Beliefs (FAB) scores were observed. Similar reductions in catastrophizing in each group were observed at each time point. At 6 months, the overall data pattern suggested no long-term difference between groups. Hence, long term bicycle riding can play a significent role in the reduction of the severe behavioural affects of chronic low back pain. (Marshall PW, Kennedy S, Brooks C, Lonsdale C. (2013) Pilates exercise or stationary cycling for chronic nonspecific low back pain: does it matter? a randomized controlled trial with 6-month follow-up.Spine;38(15):E952-9)
Frequently asked questions
Pulling up?
Mr. Krause,
This is a question about working and strengthening the muscle(s) used during the recovery phase, 180 – 360 deg (approx). Last season I began slowly working on "pulling up" during the up stroke of the pedal, with the thinking that (being 56 years old) I need everything I can get to stay with the pack during a ride. At the end of the season there was tremendous improvement, and I found general use for this little bit extra during seated climbs and sprints.
Recently, I attended a 3 day bike fitting seminar, know as SICI, during which our leader who is a PhD said it wasn't necessarily a good idea to work these muscles so much, and I could possibly cause problems if I continued. I did not argue with his position.
I've never had any problems with pain in any part of my leg(s) last cycling season. This winter (I'm in New England) I did leg presses at the gym and hit 500 lbs, leg extensions at 150lbs, and leg curls at 110lbs, which is not winning me any prizes, but I'm not embarrassed at the end of a sprint. Also, I'm not a maniac when it comes to workouts, and I generally don't mind some pain, but if I'm not getting paid to experience it, then I'd prefer not.
Could I please get your thoughts on this? My concern is not to cause a problem now, or later.
Sincerely,
G
Answer
Hi G,
The general wisdom is that you shouldn't pull up as you over-activate your hip flexor which can reduce blood flow into and out of the legs, alter lumbo-pelvic kinematics and create low back pain. As pedal force increases, the 'negative effective force' increases during the 'upstroke' phase (Blake et al 2012). In fact, Korff et al (2007) demonstrated reductions in gross efficiency when subjects were asked to 'pull up'. Furthermore, as power requirements increase, the time the knee spends in extension also increases (Elmer et al 2011) which would mitigate against pulling up. However, it sounds like you are using your hamstring muscles for the task. Elmer et al (2011) demonstrated increased knee flexor power as power requirements increased, effectively making the hamstrings a knee extensor.
Hence, the pulling up is fraut with danger if you have weak hamstrings or tight hip flexors (iliopsoas). Your hamies certainly don't sound weak with the leg curls and judging by your leg press you aren't lacking power in the quads either. Hence I don't think that the loading relative to your overall strength would lead to injury, except after a period of illness or inactivity that has led to deconditioning.
You also state you feel more power. The pulling up is important for uphill peddling and sprinting. However, the hamstring is particularly important in the downward extension phase where it is contracting and lengthening at the same time. Hence the elasticity and pliability of your hamies and their contractile recovery are important. Since you are going uphill I assume your cadence is a little slower and hence the transition phase (lengthening -> shortening) takes longer. Importantly, you should remain seated as much as possible. There is a form of strength training where athletes deliberately use high gears and even fixed gears to develop power. This differs to sprinting where power (P=Force x Velocity) is developed through the speed of muscle contraction and high cadence pedalling. Hence you may need to stretch and/or massage your hamies. But don't overstrech them and always stretch with a slightly bent knee (20 -30 degrees - bending forward at the hip) whereby you should feel the muscle pull in the middle of the hamie bulk and not at the tendons. Moreover, by bending the knee, you reduce the risk of over-stretching you sciatic nerve.
When you aren't powering uphill or sprinting you should focus on continuing to spin the Pedals as evenly as possible, even if this means on occasions using the hip flexors in the 'dead' upward phase of pedalling.
More importantly, as we age, strength training becomes very important for the prevention of sarcopenia (loss of muscle mass)
I hope that this helps
cheers
Martin
Uphill Training
- my personal preference for time poor cyclists is to do quality hill training, whereby strength and endurnace can be trained simultaneously with efficient pedalling technique
- as gradients increase to over 8% gross efficiency is lower as cadence decreases. This is compensated for by more efficient distribution of muscular force across the downward pedal cycle (Arkesteijn et al 2013, Med Sc Ex Sp, 45, 5, 920 - 926)
- slow sustained climbs involve using low gears (eg 12/13 to 53) on moderate (8%) to steep (14%) incline at a very low cadence (30rpm) for a period between 5 and 7 minutes. Total time should be in the vincinity of 21minutes (eg 3 x 7 minutes) with only a short time (4-5 minutes) for recovery.
- These slow rpms should result in some 'burning' in the leg muscles, the recognition of left - right strength deficits as well as appreciation of the upper body and core abdominal - back muscles to the overall development of synergistic strength. Hence, the cyclist should remain seated for these repeats.
- Remaining seated should mean that you are engaging the hamstring muscles to work in a synergy with the gluteal muscles and more obvious quadriceps. Importantly, if the the hamstrings are engaged correctly, there should be no sense of strain behind the knee cap (patella) or knee joint, but rather it should be felt purely within the muscle.
- Ideally, these repeates are accompanied by core stability exercises (eg 'the plank') and upper body exercises (eg japanese push-ups). Whilst doing these exercises, visualise yourself going uphill, for transference of learning to take place at a cognitive subconscious level.
- Naturally, this isn't for the faint-hearted nor should these be done in the racing season. 48-72 hours recovery between sessions is recommended. This recovery may take the form of cycling but with low gears and preferably flat.
- The program should last 4 weeks and should be done after a 6-12 week preparatory phase of endurance/base conditioning cycling.
- After these slow sustained efforts and the basic preparatory phase, a 4 week period of pre-competition hill climbing should be undertaken 2 times per week. a rest day should preceed the first session. These efforts involve finding a hill and dividing it into halves. Where the first half is at a moderate cadence of 60-70rpm and/or 70-75% HRmax, the next half involves changing down 2 gears engaging a high cadence and high power output not exceeding 85% HRmax on 1 day per week - always stay seated if possible. 48 - 72 hours later find the same hill and divide it into thirds. First third at 75% HRmax seated, the mid third at 85% HRmax out of the saddle, and the final third 90% HRmax in week 1, 95% HRmax in week 2-3 all out effort by week 4. Generally 4- 7repeats with sessions lasting between 20 and 40minutes in total. Initially, fewer repeats to accustom oneself to the intensity and later fewer repeats due to the higher intensity i.e. weeks 2 and 3 having higher repeats. Rest day should follow the higher intensity sessions whereas the first efforts should be followed by a day of 2hours of rolling hills. This trains power.
- Depending on where you live, it would be a great idea to find a hill which lasts 45-60minutes and break it up into thirds and gradually increase the intensity each third (75% HRmax, 85% HRmax, 90% HRmax). Ideally, the incline is steady and around 8-14%. This trains strength-endurance.
- At least 1 day a week should be a long ride of 3-4hours for endurance - here some high cadence (80 - 100rpm) should probably also be employed even if it is only on the flat or downhill..
- Alternate, hill training can include changes in the variables 'incline', 'cadence', 'gear ratios', 'seated vs standing' using a fartlek methodolgy whereby recovery rate is trained through 60secs on: 60secs off for a period of 20-40minutes
- If training at altitude, there are 2 philosophies - train high : sleep low and the reverese. The concept of sleeping low is based on better hormonal recovery at night after intense training. The methodology of sleeping high and training low, is that it is easier to develop more strength during training due to less strain from lack of oxygen. Then by sleeping high, the body adapts and increases it's haemoglobin carrying capacity for oxygen, which then improves endurance. With a good endurance base, strength can be developed without risking over training and injury.
Central and Peripheral Fatigue and Central Motor Drive (CMD) as it relates to altitude training
Peripheral fatigue comprises changes in the metabolic milieu of the working muscles whereas central fatigue comprises of a failure by the central nervous system (CNS) to 'drive' the motor neurons, commonly referred to as a reduction in central motor drive (CMD). It is thought that the feedback from type III and type IV nerve receptors in muscles, about the peripheral metabolic milieu, is critical to CMD. However, in an interesting review, Markus Amann (2011, Med Sc Sp Ex, 43, 11, 2039-2045) argues that, when there is a direct threat to the CNS, as a result of adverse physiological circumstances, such as hypoglycaemia, mental stress/fatigue, or in the presence of extreme environmental influences, such as heat or severe hypoxia, these factors play a much greater influence than peripheral metabolic milieu. In the case of hypoxia, the exercising terminating threshold can be up to 30% less, when exhaustion becomes exercising limiting. Hence, when training at altitude, factors such as hypoxia and the other 'threats to the CNS' should be taken into consideration, epsecially when it comes to team training camps, where many influences out of the athletes control come into play.
Muscle coordination within and between limbs
Blake et al (2012) demonstrated the importance of intra-limb coordination for the development of peak sequential power from knee to hip to ankle, and the relience on multiple muscles to produce torque. Furthermore, variations in power output at TDC and BDC are suggestive of the influence of each limb on each other. The video at the beginning of this page demonstrates the amount of 'play' which can occur across the pelvis, leading to inefficient and and potentially injurious actions. At back in Business Physiotherapy, we use a very comprehensive approach to treating the spine and pelvis, whereby, even improvements in areas of the body not traditionally associated with leg dynamics, such as ribcage biomechanics, will improve power output. Treatment of pelvic asymmetries and low back conditions can frequently relieve recalcitrant chronic knee pain. These issues have been addressed elsewhere on this website.
Game theory and the curious concept of immune - muscle synergies
It is interesting to speculate that both the novice and elite orienteer are more susceptible to infection because they both have more rigid motor patterns, are less inclined to take risk and therefore are less likely to be confronted with error generating patterns of correctability? The assumption underlying game theory is that there is a resource over which there is a conflict.
A factor which the orienteering paradigm partially defines, and identifies, is the freedom of thought and movement. That fact that many investigations are looking at exercise and the immune system, using captive animals, might be problematic in itself. These animals are in a predictable environment, which for this purpose is ethical, reductionist and conforms with rigorous scientific research protocols, which is practical, as it reduces the number of 'dependent' and 'independent' variables. However, from an evolutionary point of view, and from the perspective of the example used here, where in orienteering, more may be learnt and fine-tuned, in an unpredictable variable environment, whereby the use of caged animals may be limiting the validity of the paradigm (deterministic chaos) which is under investigation. Moreover, in terms of cognition, confinement is a stressor. Moreover, the calmness and clarity of thought needed in the freedom of orienteering (running and navigating) through the landscape can be considered the antithesis of the caged animal.
Our ancestors were striving for energy security through appeasement, subjugation or dominance over other life forms, both internally and externally. In this sense, the 'freedom to navigate and move' paradigm, suggests that our evolutionary forefathers, used their muscles, to propel themselves out of Africa, for 'greener gut biome pastures'. Therefore, maintaining good health in new and potentially 'dangerous environments', involved an unpredictable deterministic variable. As it pertains to cognition, it is an intriguing idea that a motor-sensory curiosity, drove a mechanism, which simultaneously caused muscle hypertrophy, and endowed the muscles with enhanced glutamine and cytokine concentrations, which were needed, if an immune response to pathogens was encountered. Additionally, it is conceivable, that the gut biome was driven by the laws of thermodynamics, in the search for energy. Subsequently, the gut-environment interface provided the interaction needed, to both provide energy for movement, allowed the immune system the encounters needed to flourish, and achieved the abundance of gut species required, in a variety of situations, from feast to famine. Therefore, the gut - muscle - brain cognitive pathway, was driven at a conscious and subconscious level to meet the demands of thermogenesis and immune competency, resulting in collective gain amongst all cells which required energy. Akin to Zen Buddhist philosophy, suggestions are, that the constant sensation of mild hunger, drives the clarity of thought needed, to strive for all encompassing meaning and ultimate enlightenment. Yoga, using animal poses, to stretch muscles and enhance breathing, whilst calming the cognitive state, may be based on the same primeval paradigm, of what is good for muscles is also good for the gut and brain.
The 'dove and hawk' paradigm in game theory is interesting. If a hawk and dove compete over the same resource, direct open conflict of the dove with the hawk would be suicidal. Hence, one would think it's better to be a hawk. However, what if two hawks meet, there will be only one winner and potentially both could be injured for the same net gain. Whereas, if two doves meet, they may postulate and gesticulate and eventually one may retreat or they may even share the resource, with the less risk of injury. Hence, the stakes for the Hawk are much higher. Humans have always been in conflict for resources. Does this mean that the immune system is tuned by adaptation to 'individual' resource dominance? Or, as people developed and evolved as tribes, into communities did our immune systems come to share pathogens, which may have entailed 'herd immunity' but at the same time, gave humans a lack of species variability, making them more vulnerable to 'novel' pathogens? Even more intriguing, is the notion that within a group of similar, the weakest may be driven out, seeking new resources, which in this case means new environments and new pathogens, invoking an 'immune enriching' response?
Continuing with the 'hawk dove' analogy, if we have too many hawks in a population, the consequences are constant conflict. If too many doves, constant compromise. In fact, a single dove in a hawk environment might be totally ignored, as might a single hawk in a dove environment. Thus, the apparent weaker species could be at an advantage when it's the outlier? To carry this further, we could postulate that the hawk is more prone to injury. Does this mean that in humans, the more dominant 'aggressive' individuals have an immune system which is more in demand for potential or actual injury? Whereas, the postulating dove, activates it's immune system minimally when it's performing some form of escape or deception? Interestingly, most animal conflict encounters, go out of their way to avoid injuries to themselves. Regardless, it's likely, that in both 'stressful' situations, the immune and musculoskeletal systems are activated, in a proportionately preparatory manner. Thus, the stronger the variability in the individual, the more proportional, and therefore robust their immune response.
Needless-to-say, a balance between hawks and doves needs to be established for mutual gain to take effect. In terms of the human genome, it's very underwhelming, yet it has evolved to a complex multi-cellular life form, still totally dependent upon trillions of unicellular life forms within the individual gut biome. This flies in the face of Darwinian evolution and the survival of the fittest, in that simple life forms keep complex life forms alive. Even, within the gut biome, there appears to be a symbiotic relationship with parasites and a healthy immune system (i.e. parasitic exposure appears to reduce auto-immune disease such as allergies). A 'simple' multitude (gut biome) balanced with a 'complex' minority (human DNA). Humans eat to feed their gut, their gut breaks down energy from food, which in turns propels the human, with the energy gained, to find more food. A net win win situation using the gut-brain-muscle axis.
In terms of muscles, a simplistic view, (using this analogy) would be that the glycolytic (fast twitch) muscles are the 'hawks', whereas the oxidative (slow twitch) muscles are the 'doves'. The glycolytic muscle fibres tend to be ballistic two joint (mechanical) energy straps which propel the body, whereas the oxidative endurance (metabolic energy) muscles are one joint deep muscles tending to stabilise joints and prevent injury. Therefore, the 'hawks' make the large perturbations whereas the 'doves' dampen them. Luckily, there is a muscle glycolytic fibre type which can become more oxidative (endurance power). Hence, 'hawks' which can be tempered towards a 'dove' like mentality....a balance between metabolic and mechanical energy systems, where the mechanical perturbations stimulate the metabolic-immune response.
In terms of neurophysiology, this analogy could potentially be taken further, where the sympathetic nervous system is the 'hawk' and the para sympathetic nervous system are the 'doves'. Fortunately, within the sympathetic nervous system there are self modulating ('dove' like) perturbating mechanisms. But it could be envisaged that putting a 'hawk' amongst the 'doves' wakes them up from a state of complacency. Similarly, 'staleness' is an 'existential threat' to any training regime and to any 'immune system'. Read more : Muscles, Deterministic Chaos and the Immune System
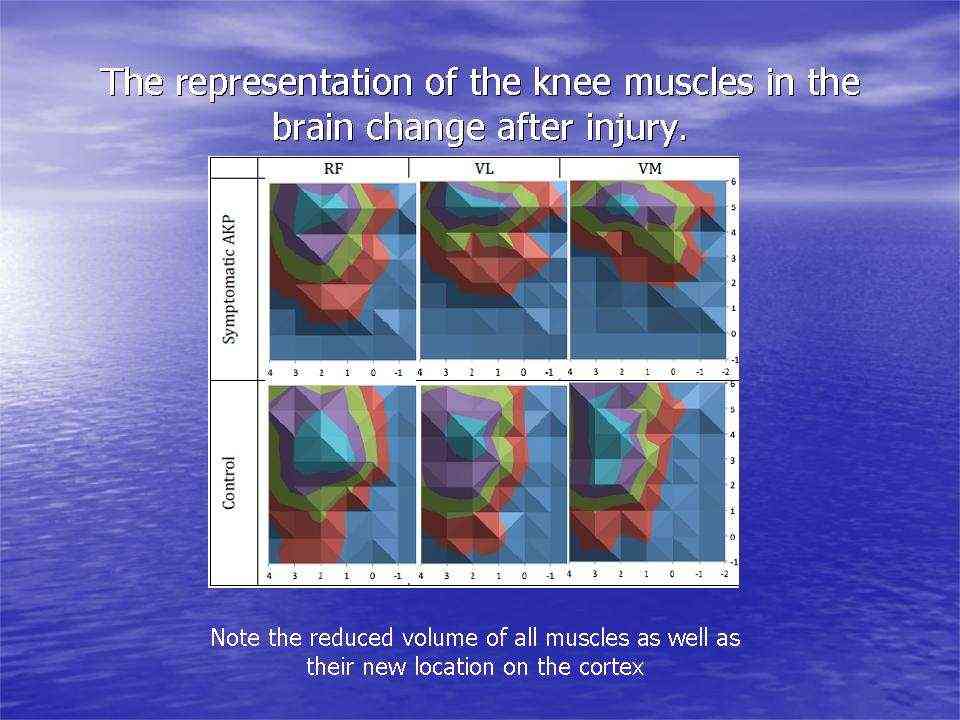
Cortical 'resource' representation, mapping, in knee muscles, after injury
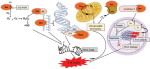
In the context of physiotherapy, a simple model of game theory might be applied to the problem of anterior knee pain. Commonly, this problem is associated with pain at the front of the knee when it is in flexion (a bent position), such as stair climbing, cycling, squats, etc. The conventional wisdom is that the patella (knee cap) is mal-aligned due to a lack of medial (VMO) muscle activity pulling the patella inwards and too much activity of the lateral thigh muscle (VL) pulling it outwards. Game theory could be applied here as the muscles compete for finite cognitive territory to represent their individual (rather than mutual) function, where ultimately neither muscle gains, both loose out to pain generating cortical representations, reducing fidelity and increasing noise in the system. However, if the muscles were to readjust for a functional outcome, they would do so through gaining a synergy of proportionally shared resources (power sharing), hereby a dominance over pain and dysfunction is gained. Physiotherapists, use functional electrical muscle stimulation (FEMS) to give added 'resources' to the VMO, which then can be used during functional exercise training. As the system adjusts, the pain improves and the gain experienced by all the muscles convinces the VL to fire with the VMO, hamstrings and gluteals in a functional synergy for stairs, cycling (see previous), squats, etc. Hereby, game theory would suggest all have won and injury has been avoided.
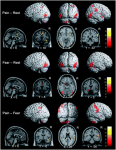
Investigations in Australia, by the Brazilian, Abrahao Baptista (2014), looking at 40-50 year old women with chronic knee pain, demonstrated the significant reduction in the volume of muscle representation in the cerebral cortices, as well as changes in the location of these muscle representations (see below). Moreover, in the context of neurons who 'fire together, wire together' it would be of utmost importance to find exercises that act as functional synergies and relate in a real or a 'virtual' manner to the environmental context in which they need to fire. This also becomes particularly important in psychological behavioural issues of 'fear avoidance' and disuse atrophy. Ideally rehabilitation stimulates 'wiring and firing together' of goal-oriented regions of the brain rather than those which are directed at fearing and avoiding movement. Therapist must instil confidence through their choices of treatment whereby they outline and hence monitor the expectations of outcome.
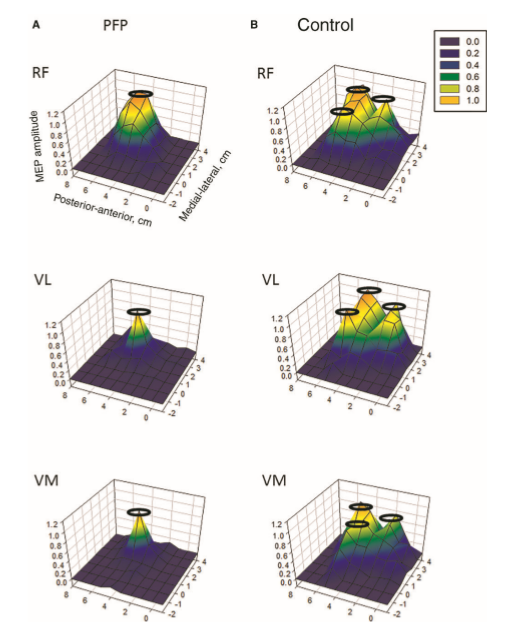
In this picture, it is clear that, the volume of the quadriceps muscle representation in the cortex of people with anterior knee pain (PFP = patello femoral pain) is dramatically reduced. Not seen here, but demonstrated in other research, is the phenomenon of pain processing altering cortical activity in various brain areas (see below).
Individuals with patellofemoral pain (PFP) had reduced map volumes and an anterior shift in the M1 representations, greater overlap of the M1 representation and a reduction in cortical peaks across all three quadriceps (RF, VL, VMO) muscles compared with controls.(Te et al 2017 Pain Medicine, pnx036, https://doi.org/10.1093/pm/pnx036). Physiotherapists use FEMS (30HZ functional electromuscular stimulation) to enhance muscle cortical representation in the brain, which improves function and in turn improves pain. FEMS stimulation readjusted cortical mapping to that seen in asymptomatic individuals. Alternatively, therapists use trans-cortical stimulation of the pre-frontal cortex to improve 'executive function' and pain.

Brain activation related to spatial discrimination of noxious stimuli is distinct from that related to perceived pain. These images are located at x = 0 mm, x = 30 mm, z = 5 mm, and y = –30 mm in standard stereotaxic space. IPL/SPL, Inferior parietal lobule/superior parietal lobule; GP/PT, globus pallidus/putamen; M1, primary motor cortex; DISCRIM., discrimination. (Oshiro et al 2007 J Neuroscience, 27, 13, 3388-3394)
Abrahao Baptista (2020) and his colleagues are conducting research into trans-cortical stimulation and vagal (para-sympathetic) nerve stimulation in the treatment of patients with COVID-19. They are basing their assumptions and working hypothesis on some positive results a rheumatologist has had with this methodology for the treatment of auto-immune inflammatory disease. COVID-19 has many of the hallmarks of an auto-immune disease, including the cytokine storm. This treatise has argued that sympathetic, rather than para-sympathetic, stimulation is necessary to add a deterministic chaotic input which perturbs the immune system into action. It would be intriguing to speculate a large perturbation between sympathetic and para-sympathetic can also activate the immune response. In this case, the illness has already set the sympathetic nervous into a heightened chaotic state, which needs additional buffering (redundancy) from the vagal nerve stimulation.
link to pins and needles, numbness in cyclists feet
Link to Pelvic - Neck dysfunction
Core Stability for cyclists
Pilates Reformer for Core Stability and Peripheral Mobility
Core Stability Training using the Swiss Ball
References
Blake OM, Champoux Y, Wakeling JM (2012). Muscle coordination patterns for efficient cycling. Med Sci Sp Exerc, 44, 5, 926 to 938.
Caldwell GE, Li L, McCole SD, Hagberg JM (1998). Pedal and crank kinetics in uphill cycling. Journal of Applied Biomechanics, 14, 245 to 259.
Caldwell GE, Hagberg JM, McCole SD, Li L (1999). Lower extremity joint moments during uphill cycling. Journal of Applied Biomechanics, 15, 166 to 181.
Cavanagh PR, Sanderson DJ (1986). The biomechanics of cycling: Studies of the pedaling mechanics of elite pursuit riders. In : Science of Cycling, Human Kinetics Books, Champaign, ch5.
Coyle EF (1995). Integration of the physiological factors determining endurance performance ability. Exerc Sports Sci Rev, 23, 25-63
Davison RCR, Swan D, Coleman D, Bird S (2000). Correlates of simulated hill climb cycling performance. Journal of Sports Sciences, 18, 105 to 110
Elmer SJ, Barratt PR, Korff T, Martin JC (2011). Joint specific power production during submaximal and maximal cycling. Med & Sc in Sp & Ex, 43, 10, 1940-1947
Fregly BJ, Zajac FE (1996) A state-space analysis of mechanical energy generation, absorption, and transfer during pedaling. Journal of Biomechanics, 29, 1, 81 to 90
Gregor RJ Fowler E (1996). Biomechanics of Cycling. In : Zachazewski JE, Magee DJ, Quillen WS. Athletic Injuries and Rehabilitation, WB Saunders Co, Philadelphia, Ch19
Jones DA, Round JM (1990). Skeletal muscle in health and disease. Manchester, Manchester University Press pp24 to 25.
Korff T, Romer LM, Mayhem I, Martin JC (2007). Effect of pedaling technique on mechanical effectiveness and efficiency in cyclists. Med Sci Sports Exerc, 39, 6, 991-995
Li l, Caldwell GE (1998) Muscle co-ordination in cycling: effect of surface incline and posture. Journal Applied Physiology, 85, 3, 927 to 934.
Lieber RL (1992). Skeletal muscle structure and function. Williams & Wilkins, Baltimore, pp41 to 42
Price D, Donne B (1997) Effect of variation in seat tube angle at different seat heights on submaximal cycling performance in man. Journal of Sports Science, 15, 395 to 402.
Rugg SG, Gregor RJ (1987) The effect of seat height on muscle lengths, velocities and moment arm lengths during cycling. Journal of Biomechanics, 20, 899
Ryschon TW, Stray-Gundersen, J (1991). The effect of body position on the energy cost of cycling. Medicine and Science in Sports and Exercise, 23, 949 to 953.
Shemmell JB, Neal RJ (1998) The kinematics of uphill, out of the saddle cycling. The North American Congress on Biomechanics, August 14 to 18, University of Waterloo, Ontario, Canada
Soderberg GL (1992). Skeletal muscle function. In : Currier DP, Nelson RM (1992) Dynamics of Human Biological Tissue, FA Davis Company, Philadelphia, Ch 3.
Stone C, Hull ML (1993). Rider/bicycle interaction loads during standing treadmill cycling. Journal of Applied Biomechanics, 9, 202 to 218.
Too D (1996) The kinematics of uphill cycling. Proceedings 9th CSB biennial conference, 184 to 185.
Too D, Lndwer GE (2000) The effect of pedal crank arm length on joint angle and power production in upright cycle ergometry. Journal of Sports Sciences, 18, 153 to 161
Van Ingen Schenau GJ (1989). From rotation to translation: constraints on multi-joint movements and the unique action of biarticular muscles. Human movement sciences, 8, 301 to 337.
Van Ingen Schenau GJ, Boots PJM, de Groot G, Snackers RJ, van Woensel WWLM (1992). The constrained control of force and position in multi-joint movements. Neuroscience, 46, 197 to 207.
Winter EM, Brown D, Roberts NKA, Brookes FBC, Swaine IL (1996). Optimized and corrected peak power output during friction-braked cycle ergometry. Journal of Sports Sciences, 14, 513 to 521.
"If everyone who lives within 5 miles of their workplace were to cycle to work just one day a week and left the car at home, nearly 5 million tons of global warming pollution would be saved every year, the equivalent of taking about a million cars off the road. "

Link to injuries relating to Pelvic Asymmetry
external non-sponsored link to Steve Hogg and his business in setting up cyclists at http://www.cyclefitcentre.com/
and see non-sponsored link to
local MTB club http://nobmob.com/
http://www.cyclingnews.com for all sorts of advice
Uploaded : 7 October 2013
Updated : 7 April 2022
 What We do
What We do What We Treat
What We Treat